 |
 |
 |
 |
CAPITULUM AND FLORET PRIMORDIA DEVELOPMENT IN PLANTS OF SUNFLOWER (Helianthus annuus L.) TREATED WITH THE MORPHACTIN CHLORFLURENOL
Luis F. Hernández
Departamento de Agronomía, Universidad Nacional del Sur, Bahía
Blanca (8000) and CIC, La Plata (1900), Argentina. ( lhernan@criba.edu.ar
)
Introduction
Many theories to explain the generation of floret pattern in capitula
of Compositae have been developed (Schwabe, 1984; Hernández, 1988;
Green, 1991; Hernández and Green, 1993; Jean, 1994). Nevertheless
the processes involved in the differentiation and pattern generation of
floret primordia are not yet well understood.
This paper reports the changes produced in the floret pattern in the
capitulum of sunflower using the morphactin Chlorflurenol (CF1: 2-Chloro-9-Hydroxy-fluorene-9-Carboxylic
acid) which main action, produced by the inhibition of IAA transport (Noodén
and Noodén, 1985), is the reduction of meristematic growth (Schneider,
1970).
Materials and Methods
Sunflower plants (Helianthus annuus L.) cv. Sunfola 68-2 were
grown under controlled environmental conditions (18 h long-day photoperiod,
500 mmol s-1 m-2 PPFD
at the canopy level, 28 °C day-night temperature) in 2 L plastic pots
containing garden soil. Plants were periodically watered and fertilized
to ensure optimal level of nutrients. Startinf at Floral Stage (FS) 4 (Marc
and Palmer, 1981), three applications of CFI were made at daily intervals,
spraying the plants with an aq. solution of 50 mg L-1 of CFI
in ethanol 20% (v/v), providing a dose of 250 mg
of CFL per plant.day-1. Control plants were similarly sprayed
with a 20% (v/v) ethanol solution.
Capitula were dissected at daily intervals, fixed and processed for
SEM [critical point dried, mounted on metal stubs and sputter coated with
gold, examined in a Cambridge S4-10 SEM at 20 kV and photographed] or light
microscopy [embedded in LR White acrylic resin, sectioned at 1-2 mm
using glass knives, the sections stained with Fluorescent Brightener 28
(Hernández and Palmer, 1988) and observed with UV light].
Results and Conclusion
The action of CFL was reflected in a complete inhibition of floret development
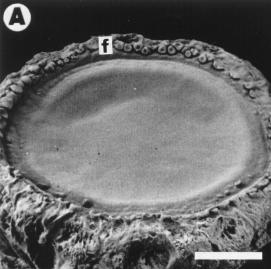
resulting in a barren area at the receptacle center (Fig. 1A-B). The morphology
of developing florets was modified and the floret corolla reduced to a
"cup shaped" structure (Fig. 1B). The orderly initiation of floret primordia
was also disrupted (Fig. 1B).
The capitula did not lose its capacity to expand (Fig 1A-B) and compared
with the controls, the receptacle sizes were only reduced at 12-15 %. This
suggests that CF1 does not arrest anticlinal cell divisions in the tunica
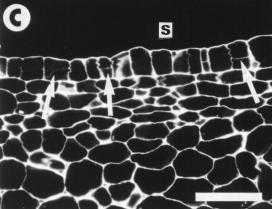
layer (Hernández, 1988). Moreover, some periclinal divisions were
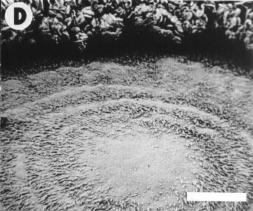
also observed in the tunica layer (Fig. 1C) resulting in the development
of hairs in the central area of some capitula (Fig. 1D). Plants allowed
to grow for a further 5-10 days, did not show any symptoms of recovery
from the CF1 -treatment and the production of floret primordia remained
suspended. Control plants (not presented in this paper) showed normal development
of floret primordia at the time of sampling.
CF1 curtailed the mitotic activity of the corpus so altering the sub
surface activity (Fig. 1C) that accompanies the formation of primordia.
Then, if applied in the transitional phase of flowering (i.e. EF4), CFL
causes inhibition of floret production and differentiation by arresting
periclinal cell divisions at the receptacle sub-surface level. Changes
observed in the floret primordia pattern can be used to further study the
way in which this pattern is naturally generated in a normal capitulum.
References
Green, P.B. 1991. Morphogenesis. In: Plant Physiology. A treatise.
Vol. 10, pp. 1-64, Academic Press.
Hernández, L.F. 1998. P.H. Thesis. Univ. of New South Wales,
Australia, 216 pp.
Hernández, L.F. and J.H. Palmer. 1988 Stain Technology,63:
190-192.
Hernandez, L.F. and P.B. Green., 1993. The plant Cell, 5:
1725-1738.
Jean, R. V. 1994. Phyllotaxis. A systemic study in plant morphogenesis.
CUP, 386 pp.
Marc, J. and J.H. Palmer. 1981. Field Crops Res., 4:
155-164.
Noodén, L.D. and S.M. Noodén 1985. Plant Physiol.
78:
263 -266.
Schneider, G. 1970. Annu. Rev. Plant Physiol., 21: 499-536.
Schwabe, W.W. 1984. Phyllotaxis. In: Positional Controls in Plant
Development, pp. 403-440, CUP.
|
|
|
|