 |
 |
 |
FLORET PRIMORDIA DIFFERENTIATION IN NATURALLY WOUNDED CAPITULA OF BORON DEFICIENT SUNFLOWER (Helianthus annuus L.) PLANTS
Luis F. Hernández
Departamento de Agronomía, Universidad Nacional del Sur, Bahía
Blanca (8000) and CIC, La Plata (1900), Argentina. (lhernan@criba.edu.ar)
Introduction
Boron deficiency during floret intiation in sunflower results in damage
of capitulum meristem and consequently in the appeareance of involucral
bracts and ray florets in different positions at the capitulum centre (Blamey,
1976, Palmer and Marc, 1982).
Palmer and Marc (1982) reported this effect after pin prick wounding
of the uncomitted receptacle surface in Floral Stage (FS) 4, 5 or 6 (Marc
and Palmer, 1981). They concluded that the wound rims could provide the
first sites for organ initials (Palmer and Marc, 1982, Hernández
and Palmer, 1988). This paper provides a detailed sequential description
of floret primordia differentiation on naturally wounded capitula from
boron-deficient sunflower plants. The implications of findings presented
here for organogenesis of the sunflower capitulum, have been extensively
discussed (Hernández, 1988; Hernández and Palmer, 1988,1991;
Hernández and Green, 1993).
Materials and Methods
Sunflower plants (Helianthus annuus L.) cv. Cargill 208A were grown
in a growth chamber (18 h long-day photoperiod, 580 m
mol s-1 m-2 PPFD at the canopy level and 28-24°C
day-night temperature) in 2 L plastic pots containing B-deficient sandy
soil. Plants were watered daily and 15 ml of a B-deficient Hoagland's solution
was added periodically to maintain optimal level of nutrients. Control
plants received an extra dosis of B (10 mg of B, as H3BO3)
every 5 days.
Starting at FS 5 (Marc and Palmer, 1981), i.e. 32-34 days after seedling
emergence (DAE), the last formed apical leaves and involucral bracts were
removed and the incipient floral meristem (receptacle) exposed. Details
of its morphology, floret differentiation and pattern formation in capitula
were then followed over time (Hernández, 1997). Once a day during
a period of 15 days, sequential replicas (Green and Linstead, 1990) of
the receptacle surface were obtained (Hernández and Green, 1993).
Sputter coated replicas were examined in a Philips 505 scanning microscope
at 10 kV and photographed.
Results
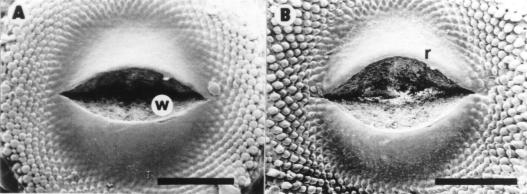
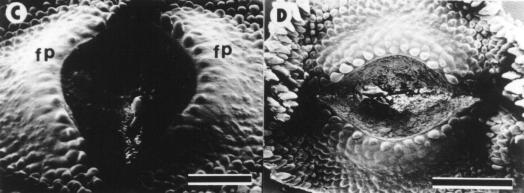
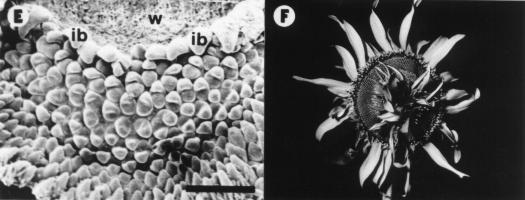
Control plants did not show morphological abnormalities and are not presented in this report. At 34 DAE, 70 % of B-deficient plants developed some sort of wounding at receptacle meritematic surface level. Only one of these plants is shown in this report (Fig. 1). At 37 DAE the wound developed into a gap 600 mm wide (Fig. 1A). At 39 DAE cell division activity in and beneath the epidermis adjacent to the wound edges (Hernández and Palmer, 1988) created an overhanging rim (Fig. 1B). At 42 DAE irregularly spaced mounds appeared on the flank of the wound rims (Fig. 1C). They rapidly developed into involucral bracts, while those formed later gave rise to ray or disc florets (Fig. 1D-E). All subsequent initials developed into disc florets (Fig. 1E). At maturity, ray and disc florets differentiated at both sides of the wound rims were normal in appearance and developed into functional structures (Fig. 1F).
References
Blarney, F.P.C. 1976. Agrochemophysica 8: 8- 1 0.
Green, P.B. and P. Linstead. 1990. Protoplasma, 158:
33-38.
Hernández, L.F. and J.H. Palmer. 1988. Am. J. Bot. 75:
1253-1261.
Hernández, L.F. 1988. Ph.D. Thesis Univ. of New South Wales,
Australia, 217 pp.
Hernández, L.F. and J.H. Palmer. 1991. Micr. Electr. Biol.
Cel., 4: 159-164.
Hernández, L.F. y P.B. Green. 1993. The Plant Cell 5:
1725-1738.
Hernández, L.F. 1997. Helia 20: 63-68.
Marc, J. and J.H. Palmer. 1981. Field Crops Res., 4:
155-164.
Palmer, J.H. and J. Marc. 1982. Plant and Cell Physiol., 23:
1401-1409.
|
|
|